How Sex Chromosomes Are More Complicated Than You Thought
Seeker (26 Jul) How Sex Chromosomes Are More Complicated Than You Thought [Video file] https://youtu.be/jhHGCvMlrb0
All females have two X chromosomes with similar genes aligned in a similar fashion. A pair of X chromosomes can function as a normal homologous pair. Like the autosomes, they can entwine with their homologous genes aligned in such a way that they can exchange substantial lengths of homologous DNA during the process of egg cell formation.
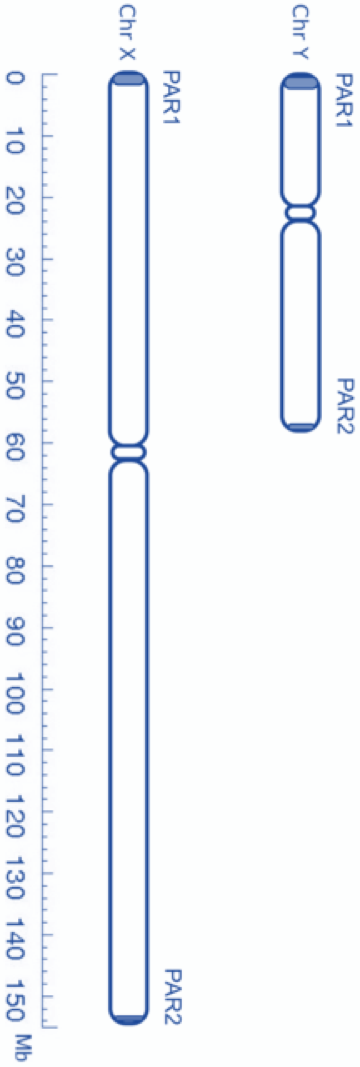
The X chromosome contains 800 - 900 genes whereas the Y chromosome is variously reported to contain only 50 to 60 genes. With such disparity in base pair and gene number, it is not surprising to learn that the X and Y chromosomes are mismatched along much of their lengths.
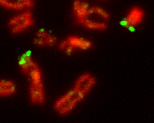
There is not much homology between the X and the Y chromosome. However, there is a bit of homology near both ends where they contain some of the same genes. We call these “matching” ends of the X and Y chromosomes the pseudoautosomal regions. Figure 2 is a micrograph of an X and Y chromosome pair stained with green fluorescent dye to show their pseudoautosomal ends. With some contortion, it is possible for limited crossover to occur between the X and Y in these pseudo-autosomal regions. This is fortunate because they are not exempt from the requirement for at least one cross-over to occur before meiosis can proceed and sperm or egg cells can be produced.
Their awkward pairing notwithstanding, these two chromosomes play an important role in human development. They are the sex chromosomes.
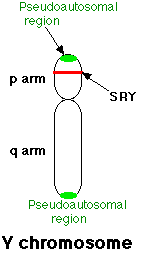
Despite its unassuming appearance, the Y chromosome contains the SRY gene that alone determines that a fertilized egg will develop into a male.
Early in the development process, the SRY gene starts a cascade of gene expression by activating genes on other chromosomes that are necessary for male development. Those genes then activate other male developmental genes and so on. For example, the SRY gene activates a gene called SOX9 that is locate on chromosome 17. Both males and females carry this gene on chromosome 17 but it can only be activated by the SRY gene. There is no SRY gene on the X chromosome. Without the SRY gene, this cascade will not start. If the SRY gene is missing or impaired, the fertilized egg will develop as a female even if there is a Y chromosome present.
All males have one X chromosome received from their mother. The unpaired genes on that chromosome are sufficient for all the the functions they control. Unlike genes residing on the paired autosomal (and paired X) chromosomes, there is no provision for masking of a deleterious recessive gene by a corresponding dominant gene that may code for a normal trait.
This unpaired state guarantees that which ever version of an allele that is present on the Y chromosome will be expressed. In most cases this is not a problem. However there is an assortment of genes on the X chromosome that may exist in some deleterious state though they are recessive. It would be rare for a female to have a copy of this allele on both of her X chromosomes. In most cases, the harmful version is masked by the presence of a more benign dominant version. However, if a mother passes the X chromosome with the harmful, recessive allele to her son, it will be expressed as there is no chance for masking it. Familiar conditions that can arise from such X linked recessive alleles are hemophilia and color blindness and Duchenne muscular dystrophy.
The mismatch of the X and Y chromosomes also results in a quirk in the expression of genes on the X chromosome in females. Since the male has only one X, over time cells have evolved to need only one copy of each X linked gene. This leads to a potential imbalance of X chromosome gene expression between male and females. To remedy this dilemma, very early in the development of the female embryo, one X chromosome in each cell is deactivated. This is accomplished by compacting the superfluous X chromosome in such a way that it cannot be unwound and made accessible for transcription to mRNA. This process is called lyonization and the resulting deactivated X chromosomal material is called heterochromatin. The deactivated X chromosome of each cell is sometimes called a Barr body.
Once an X chromosome is deactivated, it is deactivated for the life of the organism and remains inactive in all descendant cells in the organism.
The deactivation of one X chromosome occurs randomly in each cell so that the cells of a female are a mosaic of their maternal and paternal X chromosomes. A classic example of this is the Calico cat. Each colored patch on a calico cat has a different X chromosome turned on. This explains why calico cats are rarely male as they only the same X chromosome in each cell. A less well known example is women who have some patches of skin that are missing sweat glands.
An excellent, more detailed discussion of X chromosome deactivation can be found at https://www.nature.com/scitable/topicpage/x-chromosome-x-inactivation-323

Figure 1 By Ashwin Kelkar , Vivek Thakur , Ramakrishna Ramaswamy , Deepti Deobagkar [CC BY 2.5 (http://creativecommons.org/licenses/by/2.5)], via Wikimedia Commons
Figure 2 By Steffen Dietzel, Dietzel65 (Own work) [CC BY 3.0 (http://creativecommons.org/licenses/by/3.0)], via Wikimedia Commons
Figure 3 By John W. Kimball [CC BY 3.0 (http://creativecommons.org/licenses/by/3.0)], via Wikimedia Commons
Figure 4 By Shakko (Own work) [CC BY-SA 4.0 (http://creativecommons.org/licenses/by-sa/4.0)], via Wikimedia Commons
Figure 5 By Britchi Mirela (Own work) [CC BY-SA 3.0 (http://creativecommons.org/licenses/by-sa/3.0)], via Wikimedia Commons